Gut Balance
Balanced
How your pet's gut community compares to a healthy baseline. See how your pet compares with a healthy baseline.


Science-backed wellness solutions, developed and analyzed by Canadian veterinary scientists.
Test to understand. Support with daily probiotics. Reward with clean treats.

At-home microbiome and DNA testing for gut, skin, oral, and genetic health insights.
Shop Test Kits →
Daily probiotic support to help maintain healthy digestion and gut balance.
Shop Probiotics →
Plant-based, soft treats made for training, daily rewards, and sensitive dogs.
Shop Treats →The Missing Picture

Your Pet's Report
Every test comes with a clear digital report. No jargon, no guesswork. Just clear insights about your pet and practical next steps.
View a Sample ReportGut Balance
Balanced
How your pet's gut community compares to a healthy baseline. See how your pet compares with a healthy baseline.
Key Findings
What stands out
See which bacteria are below or above the healthy range. See which bacteria may need more support.
Care Plan
What to do next
Diet, supplements, routine, and what to track. Diet, supplements, routine, and what to track.
Track Over Time
Retest ready
Compare results between tests to see what's improving. Compare your baseline after a consistent care period.
Sample results shown. Your pet's report will reflect their own microbiome pattern.

Plant-based bites with prebiotic fiber (inulin & FOS).

5 billion CFU with patented strains + prebiotic inulin.
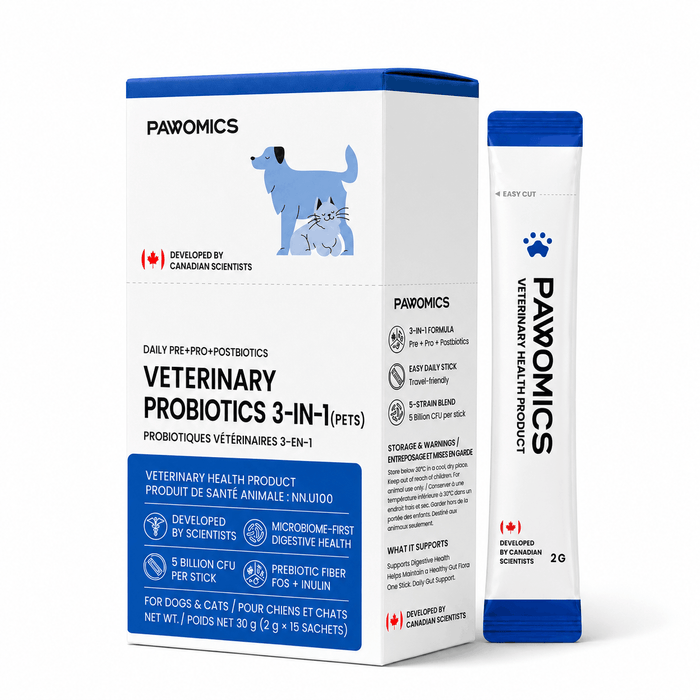
Pre + pro + postbiotic. 5 Billion CFU per sealed stick with heat-killed postbiotics.
Our dog had been scratching a lot and we couldn’t really figure out why. This test helped us understand what might be going on with his skin. The results were easy to follow.

We tested Daisy for hidden allergies with this kit, and it was way easier than we expected. It’s such an easy way to understand your pet’s health better.

You can actually test your pet’s gut health right from home with Pawomics. The instructions are extremely clear, and It provides a personalized report on gut health and diet in just weeks.

Take our quick quiz to explore your pet’s gut signals, stool patterns, and daily care needs.
Practical guides for microbiome testing, gut health, and everyday care.

Yes, some dogs can take probiotics for diarrhea, and they may help in mild or predictable cases such as ...

Choose a probiotic when your dog is generally well and you want simple daily digestive support. Choose ...

A dog’s age in human years cannot be calculated accurately by multiplying by seven. Dogs mature much...



